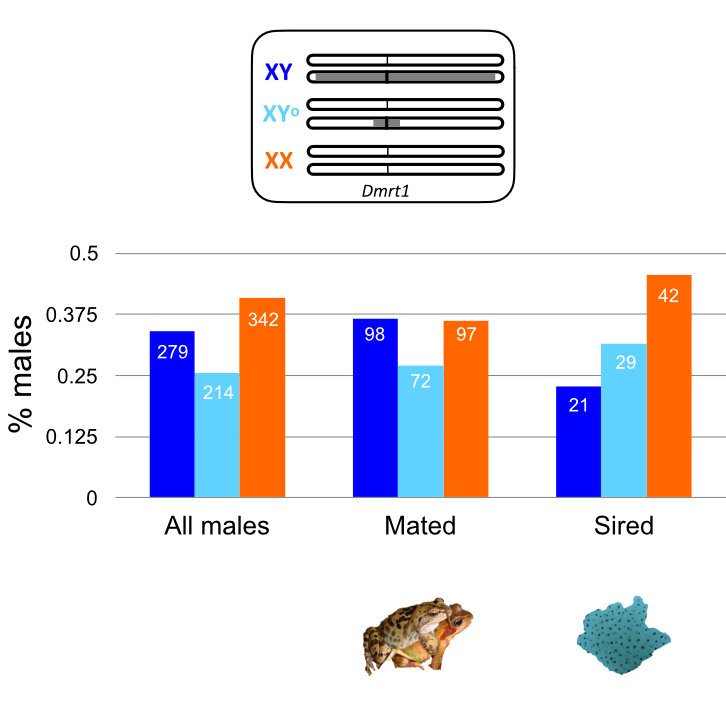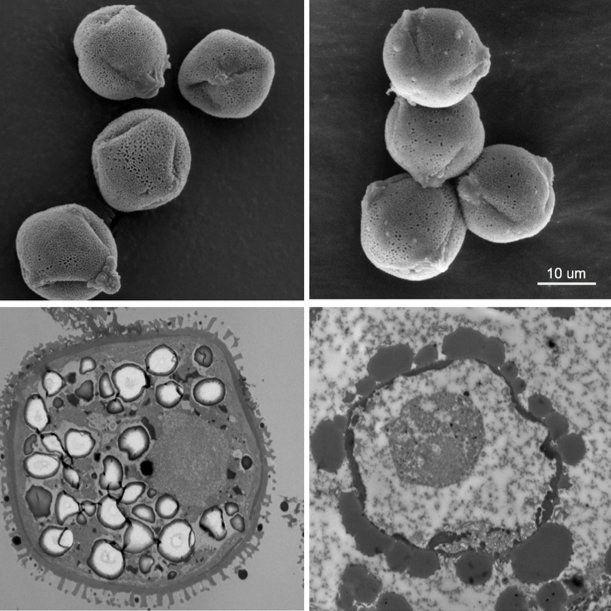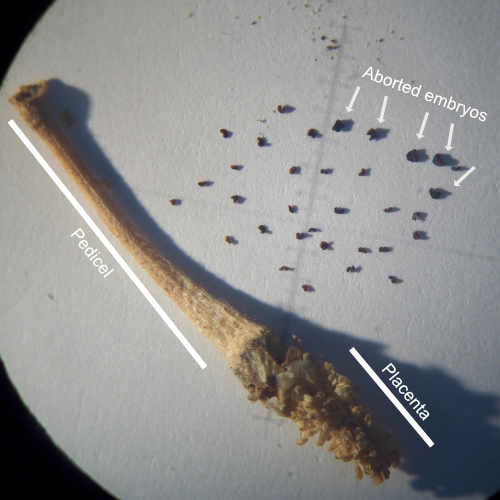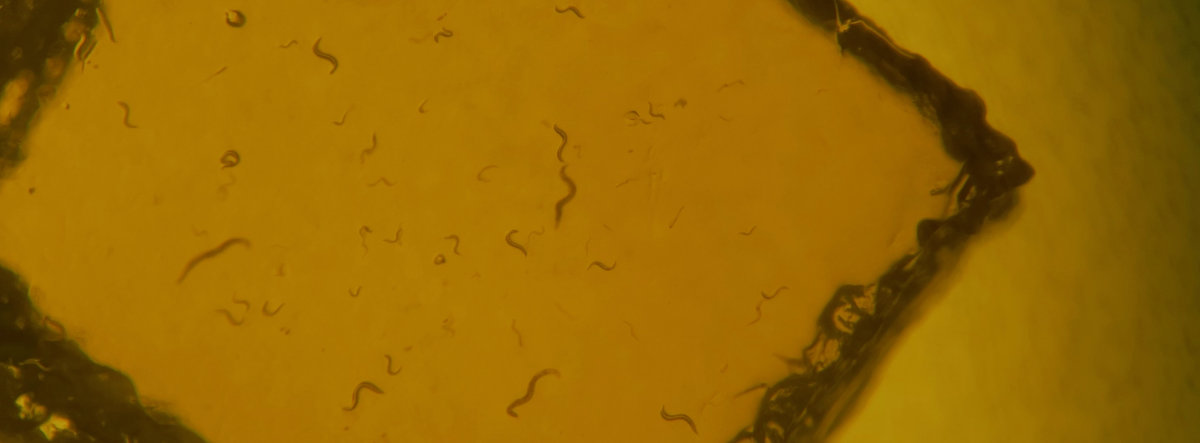






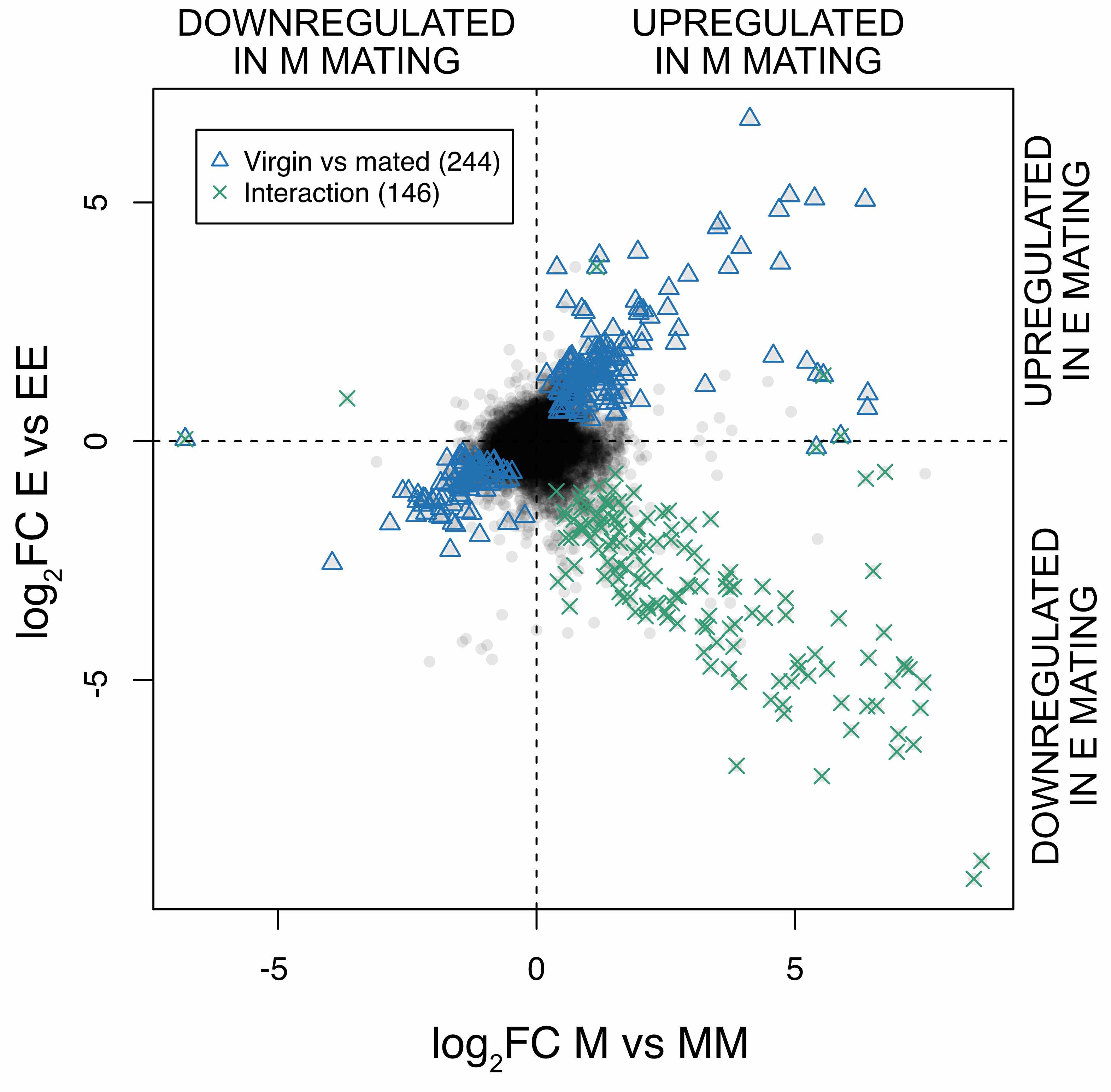
Reproductive tissue mating response after experimental sexual selection
Experimentally elevated sexual selection increases male manipulation of the female post-mating response, but also makes females poised in readiness for mating and more resistant to male manipulation.