











Double Y dosage reduces fertility, but not inviability in Mercurialis annua
Plants with 2 Y chromosomes suffer infertility, but not inviability, perhaps because XX individuals that produced male flowers had an advantage in establishing populations in the past. Or YY plants have abnormal gene dosage.
The continuous growth of plants is based on embryonic tissue they contain. Unlike animals, they make developmental pathway choices as they grow, through the differentiation of pluripotent cells into the different parts of the plant. This regeneration potential allows plants to grow from cuttings and reproduce clonally. When combined with the limited differentiation between the sexes, the regeneration ability of plants makes it possible for a normal male plant to produce female flowers, occasionally or in exceptional circumstances.
Mercurialis annua is an annual herb with male and female individuals and is an interesting plant model for studying sex chromosome evolution. While it has a XY sex chromosome system, the sex chromosomes are homomorphic and of limited differentiation, making them interesting for informing early sex-chromosome evolution theory. Males can be induced to produce female flowers by spraying them with a feminising hormone, or by repeatedly cutting the top of the plant. We used both approaches to generate female flowers and studied the phenotypic effects of having 2 Y chromosomes in the resulting offspring.
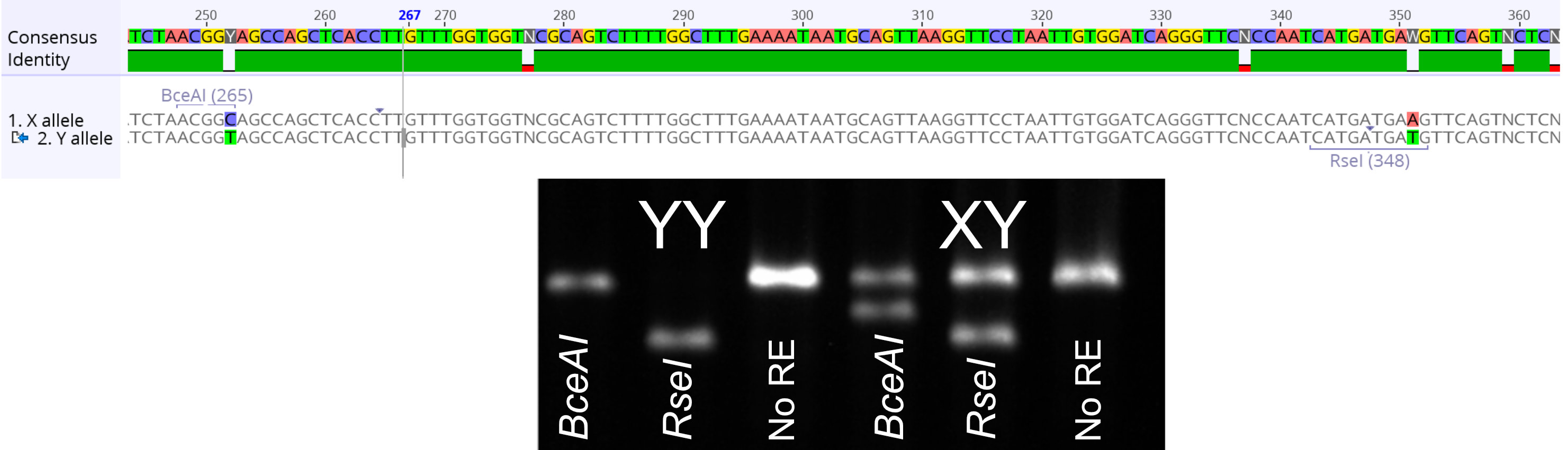
Since the female flowers originated in a XY individual, their progeny represent a XY x XY cross. We developed a molecular genetic assay to distinguish the X and Y chromosomes and genotyped the offspring. Both the genotyping and the sex ratio in the first generation suggested that YY individuals were perfectly viable. We also measured various phenotypes such as height and flower mass and did not find any major differences between XY and YY males.
We might expect the XY and YY males to differ because the Y chromosome is subject to opposing evolutionary forces. On one hand it is expected to accumulate male beneficial mutations, because the genes on the Y are always present in males. We might therefore expect a super-male phenotype from YY individuals. On the other hand, the Y has degenerated somewhat, which might result in lower fitness of YY individuals, if some genes on the Y are not working well, and a X copy of a gene is required to compensate for the degeneration of Y copies. In addition, there is an unusual gene dosage of YY plants whose ratios of Y: X or Y : Autosome genes have not been tested by selection, since these plants are very rare in nature. Like most untested changes to something that works, these dosage effects would likely work less well than the progenitor state, and result in lower fitness.
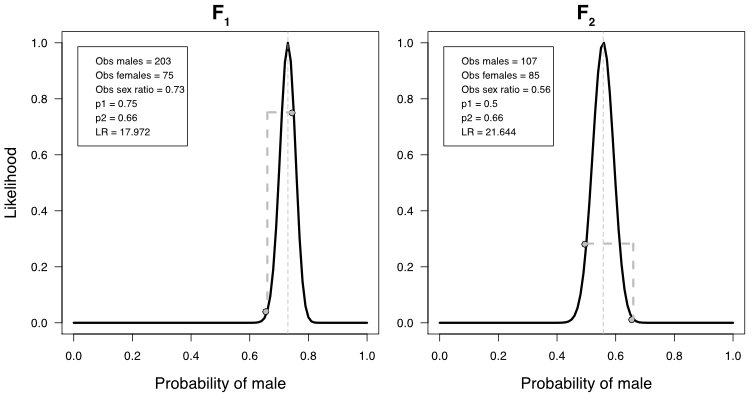
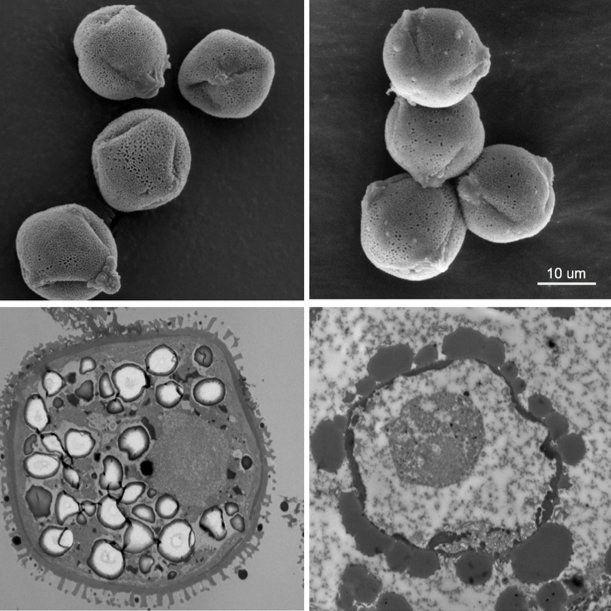
Our study of the sex ratio in the offspring of the XY and YY males and direct observations of pollen from the YY plants suggest that they are infertile, at least partially. It is surprising that fertility was affected before viability, because the Y is usually considered to be specialising in male function. For example genes on the Y improving pollen fertility are likely to be under strong selection to maintain good function, which would have selected against any mutations of genes on the Y that negatively affect pollen. Nevertheless, pollen infertility could have been caused by the dosage effects mentioned above.
We also came up with a selective explanation for our data, relating to the ability of plants to produce flowers of the opposite to their genotypic sex. This is termed sex inconstancy, and it has been found in field populations of M. annua. It is possible that the X has been under strong selection to maintain genes involved in pollen function in the recent past. For example it would be an advantage for XX individuals to occasionally produce functional male flowers, because it would allow long-distance dispersing XX individuals to establish a population on their own, without relying on long dispersing pollen for fertilisation.
In fact there is a hexaploid lineage of M. annua (in the Iberian Peninsula) in which some populations have hermaphrodite flowers, and some populations have males and hermaphrodites. This situation can be explained through a metapopulation model where hermaphrodite individuals can establish populations on their own by selfing. Once they build to a higher density they can be invaded by males, who outcompete hermaphrodites in male function. Diploid inconstant male individuals might have worked in a similar fashion in the past as they are functionally hermaphrodite even though they have separate male and female flowers. For example sex inconstancy might have conferred a fitness advantage at the end of the last ice age when new locations became available for colonisation. This would have generated strong selection to maintain pollen fertility function on the X.
More experiments will be required to understand the reasons behind the poor pollen production of YY individuals. Their initially unexpected infertility and viability has opened up new understanding in how the modular function of the plant body plan may have influenced the evolution of their sex chromosomes.
The published study is here